
你在水产养殖或技术服务中碰到过各种水质问题吗?
你在处理水质问题时碰到过效果不佳的情况吗?
你想知道是什么原因吗?
本文将从细菌及水生态系统原理出发,分析原因与处理思路,带你一步步探究水质的真相。
一、水产养殖水环境的特点及细菌的作用
天然水域生态系统中各层级的物种结构合理,数量动态稳定,生物多样性丰富,微生物、原生动物、浮游动植物、水生昆虫、节肢动物、软体动物、鱼类、甲壳类等等处于相对平衡的状态。

人工水产养殖生态系统中由于人为的因素,具有以下特点:
1、层级结构不合理
2、物质循环受阻不畅
3、生态系统脆弱
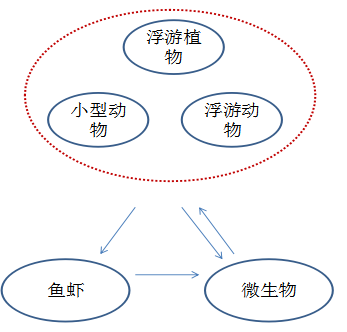
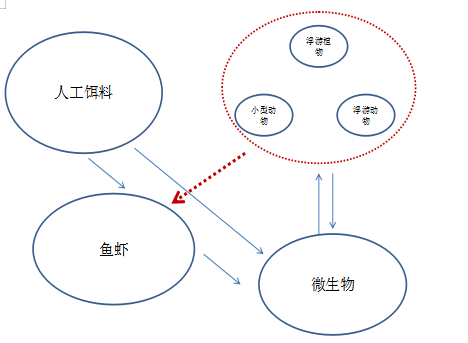
天然水域物质能量流动 养殖池塘物质能量流动
从上图可以看出,微生物在系统中的地位极其重要,同时也承担了整个养殖系统运行和稳定的核心作用,因此对池塘微生物系统功能的了解和应用是整个水质调节的基础和核心,简而言之,微生物的作用有以下四点:
1、细菌影响残饵粪便的分解
首先,细菌种类繁多,有厌氧菌、兼性厌氧菌、好氧菌等,各种细菌产生不同的酶类,这些不同细菌所产生的酶类能在不同条件下分解残饵粪便中的有机物为小分子中间物质,最后分解为二氧化碳和水。其次,部分细菌能在繁殖中将有机物转换成自身的菌体蛋白,直接间接的通过食物链传递给养殖动物。

2、细菌影响pH值
适宜的pH值是水产动物生长的必要条件,细菌的繁殖通过大量产生二氧化碳和中间代谢产物而降低环境的pH值。但同时细菌分解矿化后的营养物质促进藻类的生长,丰富的藻类通过光合作用吸收二氧化碳而增加pH值,矿化的各种矿物质又能够对水体pH值起到缓冲作用。


3、细菌影响溶氧
厌氧和兼性厌氧菌的种类和数量虽然很多,但在溶氧充足时仍然以好氧分解为主。细菌所消耗的氧气可以占到养殖水体溶氧的30%以上,但池塘底部和底泥中通常溶氧不足,这时厌氧和兼性厌氧菌的分解就非常活跃,有益厌氧细菌的繁殖不仅能够分解残饵粪便等,还能抑制有害细菌的生长。
4、细菌影响藻类
通常异养细菌分解的营养物质和生长因子能促进藻类的繁殖。但细菌中的不少种类如光合细菌类,化能自养细菌等能够与藻类竞争营养物质,抑制藻类的生长。藻周围形成的藻际环境能形成共生或敌对的复杂关系。细菌中存在大量的种类能够直接或间接裂解藻类,如已发现的粘细菌、嗜胞菌和交替单胞菌等可以直接裂解藻类。部分细菌如弧菌、假单胞菌、黄杆菌、交替假单胞菌、假交替假单胞菌等能够释放物质如蛋白、氨基酸、抗生素、羟胺等间接裂解藻类。

细菌与藻类的共生与竞争关系(张增虎等)
二、细菌在物质循环中的过程与机制
养殖水生态系统中,有机物的来源主要有:肥水或饵料未被利用的物质、排放的废物及死亡的动植物、底泥中的有机物重新悬浮。
有机物的初始分解都是由微生物所分泌的各种酶类作用,如脂肪酶、纤维素酶、蛋白酶、淀粉酶等,这些酶将各种大分子有机物质分解为小分子有机物质,小分子有机物质被吸收进一步分解、利用并转化,过程中释放出氨和二氧化碳等无机物,而无机物则被浮游植物和细菌吸收利用或通过碳氮循环途径转化。
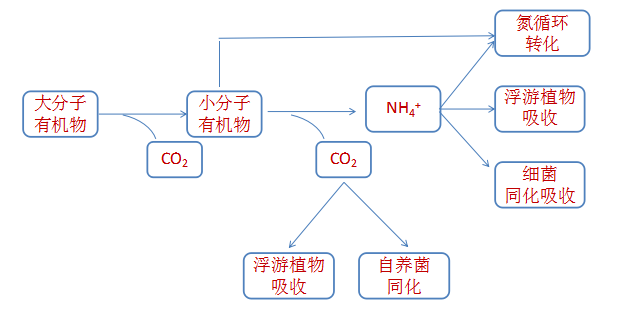
其中,氮元素的转化是整个物质转化的核心。分解有机质中的碳产物中间产物众多,大多对养殖动物无害,因此关注重点一般集中在氮的转化路径上。但碳产物和最终分解产物二氧化碳可以耦合在氮循环当中,提供能源、电子供体或受体推动氮循环的顺利进行。
由微生物参与的氮循环过程的主要转化路径可以从图表示。

氮元素的转化途径主要有六个:
途径(一):有机氮转化为氨氮,即氨化作用。
RCNH2COOH+O2—>RCH(OH)COOH+CO2+NH3
该过程中起作用的微生物种类众多,包括绝大多数的好氧菌和厌氧菌。好氧菌如常见的枯草芽孢杆菌、大肠杆菌、荧光假单胞菌、灵杆菌,厌氧菌如腐败酸菌、及兼性厌氧的酵母菌、变形杆菌等。这些微生物分泌各种蛋白酶、水解酶、氧化还原酶等酶类在细菌体外分解。通常好氧条件下进行氧化脱氮和水解脱氮,厌氧和兼性厌氧条件下进行还原脱氮、水解脱氮和脱水脱氮。

途径(二):硝化作用
NH4++1.5O2—>NO2-+2H++H2O
NO2-+0.5O2—>NO3-
该过程在好氧条件下进行,包括两个步骤,第一步先将氨氮氧化为亚硝酸盐,该过程由氨氧化细菌(Ammonia-oxidizing bacteria)和氨氧化古菌(Ammonia-oxidizing archaea)所完成。第二步将亚硝酸盐氧化为硝酸盐,由亚硝酸盐氧化菌(nitrite- oxidizing bacteria)所完成。
该途径随着氨氮的氧化,释放能量将CO2固定为自身有机物,其中每固定一个碳原子需要消耗释放能量的80%,因此硝化细菌的生长速率很低。

途径(三):反硝化作用
NO3-—> NO2-—> NO —>N2O —>N2
6NO3--+5CH3OH—>5CO2+3N2+7H2O+6OH-
该途径在厌氧条件下进行,耦合有机质作为电子供体。过程包括四个步骤,由反硝化细菌完成,但此类细菌众多,并不是每一种细菌都包括完整的四类酶。常见的有假单胞菌属、节杆菌属(Arthobacter.sp)、粪产球菌属(Alcaligenes.faecalis)等。

途径(四):硝酸盐异化还原成铵
NO3-—> NO2-—> NH4+
该过程厌氧条件下由发酵细菌(肠杆菌、气单胞菌属、梭状芽胞杆菌属等。)完成,包括两个步骤,第一步由该类细菌的异化硝酸盐还原酶将硝酸盐还原为亚硝酸盐,第二步以有机碳源作为电子供体,将亚硝酸盐还原为铵,如果有机碳源不足,第二步反应就会停止,造成亚硝酸盐的积累。

途径(五):厌氧氨氧化
NH4++NO2-—>N2+2H2O
该过程在厌氧条件下由无机碳源作为电子供体,亚硝酸盐作为电子受体将铵氧化为N2,同时将CO2固定合成有机物的过程。浮霉菌目中的厌氧氨氧化菌科如CandidatusBrocadia等细菌所完成。

途径(六):异养菌同化
α-酮戊二酸+ NH3+H+—>L-谷氨酸+H2O
丙酮酸盐+ NH3+ H+—>L-丙氨酸+ H2O
谷氨酸盐+ NH3+ATP—>谷氨酰胺+ADP+Pi
α-酮戊二酸盐+谷氨酰胺+2H++e-—>2谷氨酸盐
目前发现细菌细胞内有四种物质可以与氨氮结合,在酶和辅酶的催化下同化为有机物,这些酶包括谷氨酸脱氢酶、丙氨酸脱氢酶、谷氨酰胺合成酶、谷氨酰胺α-酮戊二酸盐转氨酶。

三、基于细菌的作用机制对常见水质问题产生原因及处理思路的分析
(一)氨氮的产生与去除路径分析
氨氮对水产动物的危害是众所周知的,主要对神经系统造成损害,损伤鳃表皮细胞。但池塘条件下的氨氮很少造成鱼虾的直接死亡,主要影响体质、摄食生长等,使得鱼虾容易发病。

通过分析氨氮的来路和去路,可以发现,氨氮产生的根本原因是高密度水体中总氮源的投入过量,在条件适宜时候,氨氮能够顺利的转化;但在某些条件下转化不畅则引起氨氮的积累。在各种氨氮的去处路径中,根据去路效率的大小可以分析有三大原因造成氨氮的积累:
1、藻相的结构不合理或藻相老化
藻类对氨氮中离子铵的去除效率极高,池塘水体在藻类丰富、数量合理、种群多样的情况下,极少有氨氮的积累。
造成藻类老化或结构不合理的因素有:1)缺磷或氮磷总量过高且氮磷比不合理,当氮磷比超过10的时候就极易暴发蓝藻。2)微量元素缺乏;水体富营养化造成藻类快速繁殖,藻类对微量元素的吸收量大且各种藻类对某些微量元素的吸收存在竞争性差异,使得吸收能力强的藻类会越来越多,吸收能力越差的藻类越来越少,最终造成藻相的单一,如爆发蓝藻、裸藻、甲藻等。
2、细菌同化吸收弱
细菌对氨氮的吸收效率变动很大,主要受营养和藻类的限制,浮游植物会与细菌竞争无机氮,限制细菌的利用。理论上细菌由于更大的比表面积,在营养条件合适时具有更大的吸收率,当补充碳源到C/N值大于10时,细菌对氨氮的吸收可以超过浮游植物。
3、氨氧化菌转化不足
氨氧化细菌和氨氧化古菌都能将氨氮转化为亚硝酸盐。但这类菌为自养菌,氧化过程中的能量80%都用于固定CO2,而且该反应必须在池塘上层的好氧条件下进行,造成转化的效率较低,但这个过程是氮元素从池塘中彻底移除出系统的必要步骤。
4、氨氮转化机制的应用分析
以上三种是氨氮去除的主要路径,厌氧氨氧化由于严格厌氧,同样存在能量利用效率低的特点。所以,在综合检测池塘的情况下可以参考的氨氮去除应用方法有以下4点:
1) 调节水体营养盐,保持N/P比小于10,补充藻类生长的微量元素。
2) 补充碳源,保持C/N比大于10。
3) 补充含氨氧化细菌和氨氧化古菌的微生态制剂。
4) 池塘增氧保持充足溶氧。
(二)亚硝酸盐的产生与去除路径分析
从亚硝酸盐的转化路径可以分析,亚硝酸盐是整个氮循环的中间产物。因此亚硝酸盐升高,总体也是由于投入氮物质的过量造成较高的平衡浓度,同时也表明转化路径受阻造成逐渐积累,具体原因如下:

1、氨氧化速率与亚硝酸氧化速率的不一致
平衡水体中两种反应保持一致,不会造成亚硝酸盐的积累。但由于各种原因引起亚硝酸氧化的速率小于氨氧化的速率就会引起积累。这些因素有:1)原始氨氧化菌与亚硝酸氧化菌的特性原因,如两种菌对底物的亲和度、氧亲和度和生长特性;2)两类菌对环境的因子的敏感性差异引起亚硝酸氧化菌的死亡或活力降低。
2、氨氧化细菌与亚硝酸氧化细菌之间空间结构被打乱
这两类菌分别参与硝化作用的两个过程,在空间上存在耦合。氨氧化菌通常形成团状结构,亚硝酸氧化菌围绕在氨氧化菌周围形成小团结构,这样在结构和功能上形成耦合,当这种结构被打破时,很容易造成亚硝酸盐的积累。
3、反硝化细菌的功能不足
硝酸盐需要经过四步过程、四种酶的作用转化为氮气,亚硝酸盐是中间产物,但很多反硝化细菌并不同时具有完整的四种酶类。有些细菌只能还原到亚硝酸盐,或有些反硝化菌在硝酸盐过高时会阻碍亚硝酸盐继续还原为氮气。
4、亚硝酸盐转化路径机制的应用分析
由上述亚硝酸盐是氮循环过程中的中间产物,主要4种微生物转化路径,且不同路径在不同的条件下进行。综合以上转化特点的去除方法有:
1) 选择具有完整功能的反硝化细菌作为微生态制剂。
2) 选择适应性强的亚硝酸盐氧化菌作为微生态制剂并用悬浮类物质做辅料。
3) 调节底部pH值为弱碱性,有利于反硝化的进行。
4) 慎用消毒剂和杀虫剂,减少环境变化对微生物的影响。
5) 添加沉性碳源与反硝化菌合剂或固定化。
四、细菌处理水质的几个观点:
1、补充细菌的安全性与可靠性问题
首先,细菌的来源应安全可靠,基因稳定性强,没有致病菌历史,没有携带可以传递的致病基因的历史等。
其次,细菌应具有较强的环境耐受能力,在不同的区域、不同品种和不同养殖模式下都能在环境中共生繁殖,并具有较强的生存能力而发挥功能作用,最为优质的菌制剂应是符合该生态系统特点的功能性土著细菌。

2、在生态功能原理的指导下,不断进行新功能菌种的开发和使用条件的应用。
水体微生物资源是一个巨大的功能宝库,其中有大量有价值的功能菌。以氮元素转化为例,在氨化、硝化、反硝化的每个阶段都有各种细菌参与,但这些具有相同功能的细菌之间并不具有近似的亲缘关系。如氨化阶段的细菌就包括变形菌门的大肠杆菌、枯草芽孢杆菌、假单胞菌,甚至包括酵母菌等,亚硝化反应的细菌包括在β-变形菌门、γ-变形菌门的多个种类,硝化反应的细菌包括在α变形菌门、硝化螺旋菌门、γ-变形菌门和d-变形菌门的多个种类。
如目前硝化反应设计主要为自养菌,繁殖世代时间长,处理水质时间久,而许多细菌、真菌、放线菌等都具有异养硝化功能如粪产碱菌(Alcaligenesfaecalis)、节杆菌(Arthrobactersp)、弯曲芽孢(Bacillus flexus)、蜡样芽孢(Bacillus cereus)、铜绿芽孢杆菌(Pseudomonas aeruginosa)等,虽然异养硝化细菌硝化效率不如自养菌,但由于异养细菌繁殖快、数量众多,因此去除氨氮亚盐效率更高。
海水中广泛存在硝酸盐异养同化细菌,筛选开发同功能淡水细菌能够增加氮的转化途径。
3、原位水处理
池塘水环境的原位处理不同于集中处理方式,不能通过微生物和添加剂以及处理条件的控制达到非常高效的处理效果。池塘中相对污染物浓度低,受多种细菌、真菌和藻类种群的相互影响,受水化学环境的制约,而且处于动态开放的系统之中,因此池塘的细菌原位处理难度更大,需要更深的理论支撑和产品支撑。首先应根据反应的各阶段准确判断细菌的处理阶段,采用合适的益生菌品种;其次采用底物亲和度更高的细菌株系,应用具有不同生态适应性的菌种,根据池塘特点调节其他指标如pH值、溶氧和C/N比等。
